La orientación magnética en los insectos sociales,
como las hormigas, avispas, abejas y termitas, ha recibido considerable
atención en los últimos años. Sin embargo, aún se conoce poco sobre los
mecanismos relacionados con la transformación de la información
magnética en alguna señal útil para la sobrevivencia de estos insectos.
Una de las hipótesis más populares es que la magnetita, un imán natural
que ha sido encontrado en abejas (Apis mellifera) y hormigas (Solenopsis sp. y Pachycondyla marginata),
puede desempeñar el papel de sensor magnético involucrado en el
fenómeno de magnetorecepción. Aquí presentamos un breve panorama de la
investigación en el área de magnetorecepción en abejas Apis mellifera
y en diferentes especies de hormigas. Incluimos nuestros estudios sobre
las propiedades magnéticas de los nanomagnetos biomineralizados por
estos insectos, realizados utilizando Resonancia Paramagnética
Electrónica (RPE), Magnetometría SQUID y Microscopía Electrónica.
Seres vivos y medio ambiente, medio ambiente y seres vivos: ¿quién
altera a quién? Los seres vivos se adaptan al medio ambiente que, por su
cuenta, se va modificando por la acción de los seres vivos. Esta
interacción que vivimos en la Tierra es una experiencia única, que no se
repite con el tiempo. Sin embargo, poco se sabe de la diversidad de los
seres vivos que habitan este planeta, o de la de aquellos que ya se
extinguieron, al igual que sobre los procesos de adaptación. Se trata de
una interacción compleja donde pequeñas alteraciones en el medio
ambiente pueden ocasionar grandes desequilibrios entre las diferentes
poblaciones, llevándolas a nuevos caminos evolutivos.Todos los seres vivos son sensibles a señales del medio ambiente, lo que posibilita, por ejemplo, la navegación y la orientación, responsables en parte de la sobrevivencia de la especie. Una de esas señales, presente desde el surgimiento de la vida en la Tierra, es el campo geomagnético. La hipótesis de que la Tierra se comporta como un enorme imán (dipolo magnético) (Fig. 1) fue presentada por primera vez en Inglaterra, en 1600, por el médico de la corte W. Gilbert. Sin embargo, desde el siglo II los chinos ya usaban los imanes para su orientación en la navegación. Los polos del campo geomagnético no siempre han estado en el mismo lugar. A veces ellos invierten su posición. Se ha descubierto que cuando esto ha sucedido, grandes grupos de seres vivos se han extinguido entre cada período de inversión, como por ejemplo, varias familias de animales diminutos, conocidos como radiolarias, que viven en la profundidad del océano (Harrison, 1968; Hays, 1971).
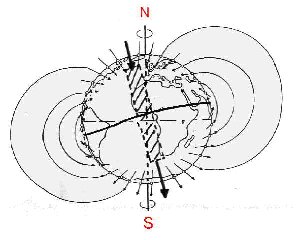
Figura 1. Comparación de las líneas de fuerza del
campo geomagnético con las de un dipolo magnético (o un imán). Nótese la
inclinación entre los ejes N-S magnético y geográfico. Las líneas
negras discontinua y continua representan a los ecuadores geográfico y
magnético, respectivamente.
A pesar de relatos bastante antiguos sobre la migración de ciertas
aves, sólo desde la segunda mitad del siglo XX se realizan estudios
sistemáticos sobre la capacidad migratoria de éstas, y en particular,
sobre su capacidad de orientarse detectando el campo geomagnético.?Pero,
¿qué significa detectar el campo geomagnético? ¿Existe algún sensor
especializado en la detección de campos magnéticos en estos animales?
Hasta hoy sólo se conoce bien la magnetotaxia, que es el mecanismo de
interacción entre este campo y algunos microorganismos. Las bacterias
magnéticas y algunos organismos unicelulares producen partículas
magnéticas suficientes para orientarlas siguiendo las líneas del campo
geomagnético. Esta es una respuesta pasiva, funcionando como una
brújula, donde la aguja gira alineándose con la dirección de este campo
(Farina et al., 1990). En el caso de los animales superiores, como
abejas, moscas, mariposas, tortugas, salamandras, salmones, atunes,
ballenas, delfines, tiburones, palomas, etc., los diferentes mecanismos
de detección del campo magnético son poco conocidos. En varias de estas
especies se han encontrado partículas de material magnético
biomineralizado, en general magnetita, que es el óxido de hierro
magnético más común en la naturaleza, con tamaños de aproximadamente
cuatro a diez millonésimos de centímetro. El mecanismo de detección del
campo magnético en estos animales (conocido como magnetorecepción [1]),
es mucho más complicado que en los microorganismos.Entre los animales superiores, los insectos constituyen la clase dominante (en número de especies y biomasa). Abejas, hormigas, avispas y termitas pertenecen al grupo de los insectos sociales y viven en colonias, formando sociedades organizadas en tres castas básicas: obreras, soldados y reinas. La orientación magnética en estos insectos ha sido estudiada (Vácha, 1997), y el insecto en el cual se han hecho más estudios, demostrando su capacidad de detectar el campo geomagnético, es la abeja Apis mellifera (Fig. 2). La presencia de magnetita en el abdomen de esta especie de abejas ha estimulado la aplicación y el desarrollo de la hipótesis ferromagnética para explicar su magnetorecepción (Gould et al., 1978).

Figura 2. Abeja Apis mellifera.
Se sabe que las abejas ejecutan una danza en la colmena, tomando
como referencia el campo gravitacional de la Tierra. Las abejas
forrajeras, cuando regresan de una exitosa búsqueda de alimento,
ejecutan una danza cuya orientación en relación a la dirección vertical
de los panales de la colmena indica, a las otras abejas, la localización
de la fuente de alimento. El ángulo entre la dirección de la danza y la
vertical indica el ángulo entre la fuente de comida y el Sol (ver
Chittka & Dornhaus, 1999). En esta danza, han sido observados
ciertos "errores" de hasta 200 a la izquierda o a la derecha
de la dirección "correcta", variando con la dirección del campo
geomagnético. Estos "errores" no son "ruidos del sistema", pues todas
las abejas, danzando en un instante dado cometen el mismo "error", tanto
en intensidad como en dirección (ver Wiltschko & Wiltschko, 1995).Por otro lado, cuando un enjambre de abejas deja la colmena original, abejas obreras de este enjambre construyen nuevos panales en la misma dirección magnética de la colmena anterior. Son necesarios campos magnéticos relativamente fuertes (» 10 veces el de la Tierra, el que en Rio de Janeiro, Brasil, es de apoximadamente 0.25 Oe) para destruir esta orientación de los panales.
Las preguntas, aún sin respuesta, son: ¿Cómo hacen esto las abejas? ¿Cuál es la naturaleza de su sensor magnético? Como ya fue mencionado, la hipótesis más probable para la magnetorecepción, en el caso de la abeja Apis mellifera, esta basada en la presencia de nanopartículas de magnetita en su abdomen. Las propiedades magnéticas de estas partículas dependen de su tamaño y forma, y pueden ser catalogadas como: multidominios magnéticos (donde la configuración total de los momentos magnéticos de cada dominio da lugar a una baja energía magnética en toda la partícula), monodominios magnéticos (caracterizadas por tener un momento magnético estable) o superparamagnéticas (magnéticamente inestables debido a la energía térmica del medio externo). En general, estas partículas tienen un comportamiento magnético diferente, ya que mientras las de mayor tamaño (multidominios y monodominios) están permanentemente magnetizadas, las menores (superparamagnéticas), con tamaños por debajo de un cierto tamaño crítico, pueden modificar su vector de magnetización y aún perder la magnetización por variaciones en la temperatura, sin que estas partículas se muevan. Así, estas últimas responden rápidamente a variaciones del campo magnético, pudiendo de esta forma, desempeñar el papel de sensor de la variaciones magnéticas del medio ambiente durante el vuelo de las abejas (Wiltschko & Wiltschko, 1995).
Nuestros resultados (aun no publicados) con Resonancia Paramagnética Electrónica confirman la presencia de partículas superparamagnéticas aisladas de magnetita, así como de agregados de las mismas en el abdomen de abejas Apis sp.
¿Y las hormigas?
Con respecto a las hormigas, el número de especies existentes en el
planeta es superior a 12,000 y en Brasil, debido al clima y a las
selvas tropicales, pueden encontrarse una gran parte de estas especies.
Todos nosotros mantenemos un contacto diario con las hormigas. Basta dar
una mirada a la cocina de nuestro apartamento, o en el patio de nuestra
casa, para que nos encontremos con estos insectos buscando alimento u
organizando el nido donde viven. En general, los hormigueros debajo de
la tierra están formados por varios túneles subterráneos en completa
oscuridad, los cuales irradian a partir de un punto central y terminan
en salidas por donde ellas llegan a la superficie. A pesar de las
diferencias de cada especie en cuanto al proceso de descubrir y
transportar alimento a la colonia, lo que ellas tienen en común es que
dejan el nido y exploran las áreas alrededor del mismo, dando vueltas en
un patrón aleatorio hasta encontrar alimento, momento en el cual
regresan al nido, marcando el camino de vuelta con feromonas
características de cada colonia. Este camino de regreso es recto en la
dirección de la salida del túnel, independientemente de lo azaroso del
camino empleado en la búsqueda de alimento (Hölldobler & Wilson,
1990). Es bastante claro que ellas deben ser sensibles a las diferentes
fuentes de información existentes en la naturaleza, como la posición del
Sol, la polarización de la luz celestial, el patrón geométrico que las
ramas de los árboles forman en el techo celeste, el paisaje del
horizonte cercano y el campo geomagnético, entre otras.Uno de los primeros estudios hechos para mostrar la sensibilidad de las hormigas a campos magnéticos, fue hecho por Kermarrec en 1981. En su laboratorio, colocó imanes intensos cerca de nidos artificiales de hormigas Acromyrmex octospinosus. Kermarrec observó consistentemente que ellas evitaban las regiones que quedaban cerca de los imanes. Este comportamiento de "repulsión magnética" es una evidencia de la sensibilidad de las hormigas a campos de fuerza alterados en su entorno.
En 1993, Anderson y Vander Meer hicieron un estudio de laboratorio con la hormiga Solenopsis invicta, conocida en EEUU como "hormiga de fuego" (fire ant) por el dolor agudo que causa su picada. El experimento consistió en lo siguiente: un nido de hormigas artificial fue colocado dentro de un artefacto que genera campo magnético, conocido como bobina de Helmholtz, para poder manipular el campo geomagnético. Fue colocada una cucaracha como alimento para las hormigas: desde el momento que una hormiga exploradora encontraba la cucaracha, el tiempo de reclutamiento y formación de columna era medido. Dos situaciones básicas fueron estudiadas: el tiempo con el campo geomagnético inalterado y el tiempo con el campo geomagnético alterado a partir de que la exploradora encontraba a la cucaracha. La alteración consistia en una inversión del componente del vector de campo geomagnético paralelo al suelo. Lo que se observó es que el tiempo en el caso del campo alterado fue el doble de aquel tiempo con campo inalterado. Esto demostró la sensibilidad de las hormigas al campo geomagnético, pero no demostro que las hormigas puedas orientarse usando el campo geomagnético.
El trabajo de Çamlitepe y Stradling, en 1995, demostró que las hormigas pueden usar la información direccional del vector de campo geomagnético como una referencia espacial para la orientación. Ellos hicieron estudios en el campo y en el laboratorio con hormigas de la especie Formica rufa. En este estudio, las hormigas fueron condicionadas a buscar alimento en la dirección Norte magnética. Cuando esta dirección fue alterada, la mayoría de las hormigas fueron a buscar alimento en la nueva dirección del Norte magnético. Esto significa que las hormigas son capaces de usar la dirección y el sentido del campo para regresar a un lugar específico.
Estos estudios realizados en diferentes especies de hormigas han servido para determinar su capacidad para sentir y usar el campo geomagnético en diversas situaciones. Sin embargo, son necesarios más estudios para verificar esta hipótesis.
Nuestras investigaciones en diferentes especies de hormigas han confirmado la presencia de material magnético en las mismas (Esquivel et al., 1999; Acosta-Avalos et al., 1999). Una especie interesante es Pachycondyla marginata (Fig. 3) que muestra un comportamiento migratorio. Ellas se encuentran al sudeste de Brasil y sólo se alimentan de termitas de la especie Neocapritermes opacus. Un análisis nuestro de las rutas anuales de migración de varias colonias muestra una preferencia para escoger rutas de migración en un eje desviado aproximadamente 12º del eje Norte-Sur magnético (resultados no publicados). Estos resultados sugieren la capacidad de esta hormiga para utilizar la información del campo geomagnético durante el proceso de migración. Al igual que con la abeja A. mellifera, se pensamos que la hipótesis ferromagnética de la magnetorecepción también podría ser aplicada a esta hormiga. Para demostrarlo aislamos nanopartículas de óxidos de hierro magnéticos de su abdomen, y usando microscopía electrónica las identificamos como partículas de magnetita o maghemita. Además del abdomen, la cabeza también puede estar involucrada en el proceso de magnetorecepción, debido a la presencia de estas nanopartículas en la misma (Acosta-Avalos et al., 1999).

Figura 3. Hormiga Pachycondyla marginata cargando una termita, su único alimento.
Medidas de la magnetización inducida, hechas a través de
magnetómetros acoplados con SQUID, en abdómenes secos y machacados de
estas hormigas, mostraron la existencia de propiedades
ferro(i)magnéticas en coexistencia con un estado superparamagnético a
temperatura ambiente (resultados no publicados). Esto puede implicar la
existencia de dos tipos diferentes de distribuciones de nanopartículas
magnéticas en el abdomen de estas hormigas. Complementando nuestro
trabajo, los estudios de RPE en estos abdómenes (Wajnberg et al., 2000)
revelaron la presencia de partículas aisladas con volúmenes magnéticos
compatibles con los que fueron medidos por microscopía electrónica, y
también de pequeños aglomerados.Una característica importante de la magnetorecepción y de los sensores magnéticos biomineralizados es que son específicos de cada especie. Aún existe un largo camino por recorrer para la comprensión total de este tipo de mecanismo, tomando en cuenta los pocos datos existentes y la enorme diversidad de especies. Aunque se están estudiando comportamientos relacionados con campos magnéticos aplicados en hormigas y abejas, es necesario que, paralelamente, se estudien las características y las propiedades magnéticas del material biomineralizado, para que modelos más adecuados sean propuestos. Siguiendo este último criterio, nuestro grupo ha investigado hormigas y abejas que viven en Brasil. Así, utilizando técnicas físicas aplicadas a estos materiales magnéticos biomineralizados, será posible generar un conjunto de resultados y contribuir a la compresión del mecanismo de percepción del campo magnético.
Notas
Para entender los mecanismos de magnetorecepción, se han postulado tres mecanismos diferentes:- la hipótesis ferromagnética, que involucra la existencia de partículas magnéticas como transductores del campo magnético.
- la modificación del mecánismo de visión, a través de la creación de radicales químicos por pares, la cual depende de la presencia de campos magnéticos.
- la inducción electromagnética, en la cual variaciones del flujo magnético pueden producir campos eléctricos y corrientes de iones. Este mecanismo ya ha sido encontrado en algunos peces eléctricos.
Bibliografía
Acosta-Avalos, D., Wajnberg, E., Oliveira, P.,
Leal, I., Farina, M. & Esquivel, D.M.S. (1999). Isolation of
magnetic nanoparticles from Pachycondyla marginata ants. J. Exp. Biol., 202: 2687-2692.
Anderson, J. & Vander Meer, R. K. (1993). Magnetic orientation in fire ant Solenopsis invicta. Naturwissenschaften, 80: 568-570.
Çamlitepe, Y. & Stradling, D. J. (1995). Wood ants orient to magnetic field. Proc. R. Soc. Lond. B, 261: 37-41.
Chittka, L. & Dornhaus, A. (1999).
Comparaciones en fisiología y evolución, y por qué las abejas pueden
hacer las cosas que hacen. Ciencia al Dia Internacional, 2 (2): http://www.ciencia.cl/CienciaAlDia/volumen2/numero2/articulos/articulo5.html
Esquivel, D. M. S., Acosta-Avalos, D., El-Jaick,
L.J., Linhares, M.P., Cunha, A.D.M., Malheiros, M.G. & Wajnberg, E.
(1999). Evidence for magnetic material in the fire ant Solenopsis sp. by Electron Paramagnetic Resonance measurements. Naturwissenschaften, 86: 30-32.
Farina, M., Lins de Barros, H. & Esquivel, D.M.S. (1990). Organismos magnetotácticos. Investigación y Ciencia, 171: 70-78.
Gould, J. L., Kirschvink, J. L. & Deffeyes, K. S. (1978). Bees have magnetic remanence. Science, 201: 1026-1028.
Harrison, C. G. A. (1968). Evolutionary processes and reversals of the Earth’s magnetic field. Nature, 217: 46-47.
Hays, J. D. (1971). Faunal extinctions and reversals of the Earth’s magnetic field. Geol. Soc. Am. Bull., 82: 2433-2447.
Hölldobler, B. & Wilson, E. O. (1990). The ants. Harvard University Press. Cambridge.
Kermarrec, A. (1981). Sensibilité à un champ magnétique artificiel et reaction d’évitement chez Acromyrmex octospinosus (Formicidae, Attini). Insec. Soc. Paris, 28: 40-46.
Vácha, M. (1997). Magnetic orientation in insects. Biologia Bratislava, 52: 629-636.
Wiltschko, R. & Wiltschko, W. (1995). Magnetic orientation in animals. Springer-Verlag, Berlin, Heidelberg.
Wajnberg, E., Acosta-Avalos, D., El-Jaick, L. J.,
Abraçado, L., Coelho, J. L. A., Bakuzis, A.F., Morais, P. C. &
Esquivel, D. M. S. (2000). Electron Paramagnetic Resonance study of the
migratory ant Pachycondyla marginata abdomens. Biophys. J., 78: 1018-1023.
No hay comentarios:
Publicar un comentario